|
Article Open access Published: 05 January 2026
HIV disrupts the lung molecular clock, leading to lung inflammation and features of emphysema
- Kingshuk Panda, Joyce Yao-chun Peng,
Maria J. Santiago, Md. Sohanur Rahman,
Sadiya Bi Shaikh, Gagandeep Kaur,
Md Imam Faizan, Khursheed Ul Islam,
Ting Wang, Stephen M. Black,
Christopher M. Evans, Irfan Rahman,
Srinivasan Chinnapaiyan, & Hoshang J. Unwalla
Communications Biology volume 9, Article number: 11 (2026) Cite this article
Abstract
In the aging HIV (human immunodeficiency virus)-infected population, comorbid diseases are important determinants of morbidity and mortality. People living with HIV (PLWH) demonstrate increased lung inflammation and incidence of chronic obstructive pulmonary disease (COPD), even after adjusting for smoking status. Disruption of the lung molecular clock has been implicated in the increased lung inflammation observed in COPD and smokers. We hypothesize that the expression of HIV TAT protein in the lungs of PLWHpromotes lung inflammation and features of emphysema due to dysregulation of lung circadian rhythm, determined by the lung molecular clock genes. We demonstrate that HIV TAT upregulates miR-126-3p in primary bronchial epithelial cells and suppresses Sirtuin 1 (SIRT1), resulting in downstream effects on core circadian genes such as BMAL1 and PER2, leading to dysregulation of the lung molecular clock. This study identifies TAT/miR-126-3p/SIRT1 axis as an important mediator of HIV-induced lung inflammation in primary human bronchial epithelial cells, SPC-TAT transgenic mice with lung-specific TAT expression, and lungs from HIV-positive donors. Using single-cell RNA sequencing of lungs from 4-month-old SP-C TAT mice, we further show that these mice already exhibit significant alterations in clock gene expression and elevated expression of proinflammatory markers in their young adult stage. Understanding the pathophysiological mechanisms by which HIV disrupts the lung molecular clock and promotes inflammation may help identify therapeutic strategies to mitigate HIV-associated COPD.
Introduction
Globally, ~39 million people are estimated to be living with human immunodeficiency virus (HIV)1. While access to combination antiretroviral therapy (cART) has increased, a cure remains elusive, as cART alone is incapable of eradicating HIV reservoirs2. In the aging, HIV-positive population, comorbid diseases are essential determinants of morbidity and mortality3. People living with HIV (PLWH) receiving effective cART tend to die from non-acquired immune deficiency syndrome (AIDS)-related comorbidities nearly a decade earlier than those without HIV4. Chronic obstructive pulmonary disease (COPD) is a major comorbidity in PLWH, with HIV infection serving as an independent risk factor. The progression and severity of COPD are further worsened by cigarette smoking5.
COPD exacerbations demonstrate a diurnal rhythm, with more frequent exacerbations occurring during the night or early morning hours, likely due to circadian dysregulation affecting mucus production and lung inflammation6,7,8. The circadian clock constitutes a nearly 24 h oscillation in gene expression, leading to changes in physiological processes, metabolism, and behavior, and thereby orchestrating the rhythmic synchronization of physiological processes9,10,11. In mammals, the circadian timing system consists of a central pacemaker clock located in the suprachiasmatic nucleus (SCN) of the brain, along with additional peripheral clocks found in most tissues throughout the body12. Although light/dark cycles are the primary zeitgebers (timing cues) for the SCN pacemaker that subsequently synchronizes peripheral clocks, feeding patterns and environmental factors can also act as potent zeitgebers, influencing the tuning or disruption of peripheral clocks13.
In the lung, the circadian rhythm regulates daily fluctuation in airway diameter, airway resistance, respiratory symptoms, mucus production, and inflammatory responses, which in turn contribute to the diurnal pattern of exacerbation frequency and occurrence14,15.
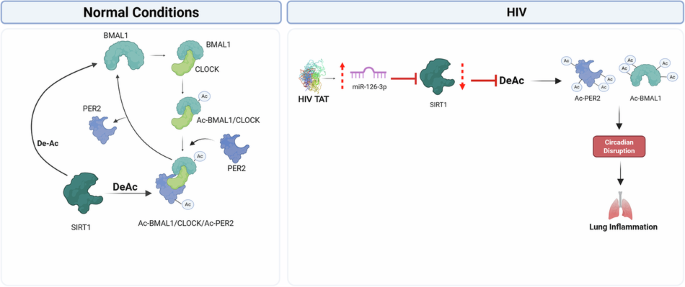
The molecular clock operates through a negative feedback loop involving various transcription factors known as clock genes. In mammals, the CLOCK:BMAL1 heterodimer promotes the expression of period (PER1-3) and cryptochrome (CRY1-2) genes. PER and CRY form phosphorylated heterodimers that translocate back to the nucleus, where they inhibit the activity of the BMAL1:CLOCK complex16,17.
Post-translational modifications including as acetylation and phosphorylation, play critical roles in regulating the activity and stability of molecular clock proteins. In the lung, clock dysfunction can lead to alterations in pulmonary physiology and contribute to disease pathologies, particularly in response to environmental stressors such as cigarette smoking14. Sirtuin 1 (SIRT1) is the primary circadian deacetylase linking core clock functions with inflammatory responses in the lung. It deacetylates BMAL1 and PER2, modulating their activity and, by extension, sustaining the circadian expression and activity of these clock proteins. The suppression of SIRT1 in airway epithelia of smokers and COPD patients is associated with a dysregulated molecular clock and consequent lung inflammation and injury10,18,19. Recent studies, including ours, show that clock dysfunction in airway epithelial cells plays an important role in pulmonary physiology and pathology, particularly in response to proinflammatory mediators such as cigarette smoking14,18.
MicroRNAs (miRNAs) play an important role in post-transcriptional regulation of genes and are often dysregulated in disease20. HIV transactivator of transcription (TAT) is a viral protein secreted by infected cells that enters bystander cells and alter gene expression independent of viral replication. In our study, we employed both whole HIV and purified Tat protein to delineate TAT-specific effect. We previously reported that HIV TAT alters the bronchial epithelial microRNAome21. Here, we identify novel miR-126-3p-mediated SIRT1 regulation as being,primarily responsible for HIV-mediated dysregulation of the lung molecular clock. These observations were validated in primary normal human bronchial epithelial cells (NHBE), SP-C TAT transgenic mice (lung-specific TAT expression), and HIV-positive human lungs. We further demonstrate that this dysregulation has downstream effects on the lung, including lung inflammation. ScRNAseq experiments on lungs from young adult SP-C TAT transgenic mice demonstrated that HIV TAT expression, resulting in circadian disruption and inflammation, becomes evident in multiple cell types at very early stages of life. The significance of our findings will be discussed.
Results
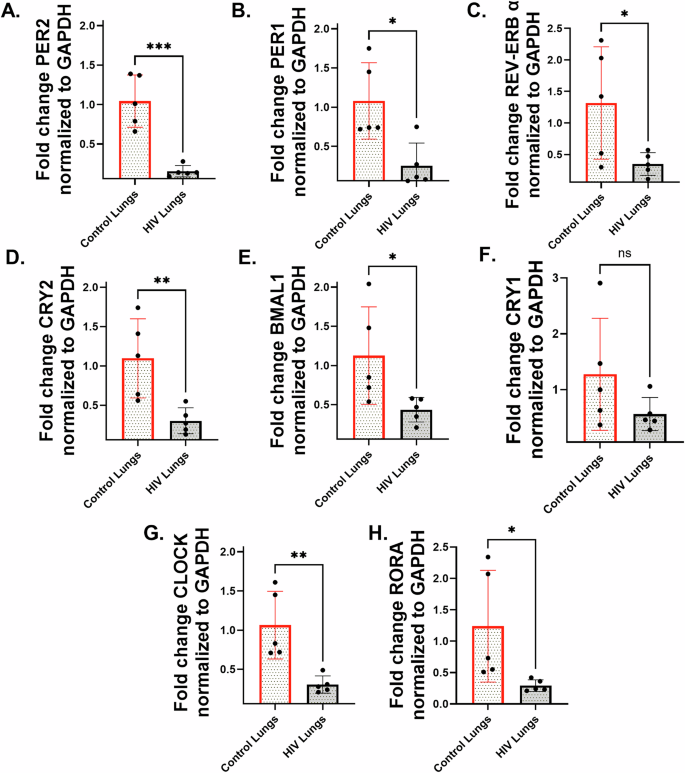
Circadian genes are suppressed in lungs from HIV-positive donors
Total RNA was quantified for expression of major circadian genes. Quantitative reverse transcription-PCR(qRT-PCR) analysis revealed a significant reduction in the expression of major circadian clock genes in HIV-positive lung tissues compared to HIV-negative controls. Specifically, there was a statistically significant downregulation of PER1, PER2, REV-ERB α, CRY2, BMAL1, CLOCK, and RORA (Fig. 1A–H). Interestingly, the expression of CRY1 also trended lower in HIV-positive paired donor lungs, although the suppression was not significant (Fig. 1G, H). While expression of circadian genes in non-HIV control lungs displayed greater inter-individual variability, consistent with known sensitivity of peripheral circadian rhythms to lifestyle and physiological factors, HIV-positive lung samples exhibited a more uniform suppression of circadian genes reflecting the dominant influence of viral proteins such as Tat on circadian regulation, which may override inter-individual variability.
To determine if our HIV positive samples demonstrate Tat expression in the lungs we performed an Immunohistochemistry (IHC) analysis using a HIV Tat-specific antibody. Tat immunoreactivity was localized to the alveolar and airway epithelium and was accompanies by infiltration of inflammatory cells only in lung sections from HIV positive donors. No detectable Tat staining was observed in HIV-negative lung tissue (Supplementary Fig. 1). HIV-positive lungs show approximately sixfold increase in staining intensity compared to HIV-negative controls.
HIV TAT dysregulates the expression of BMAL1 and PER2 in NHBE ALI cultures
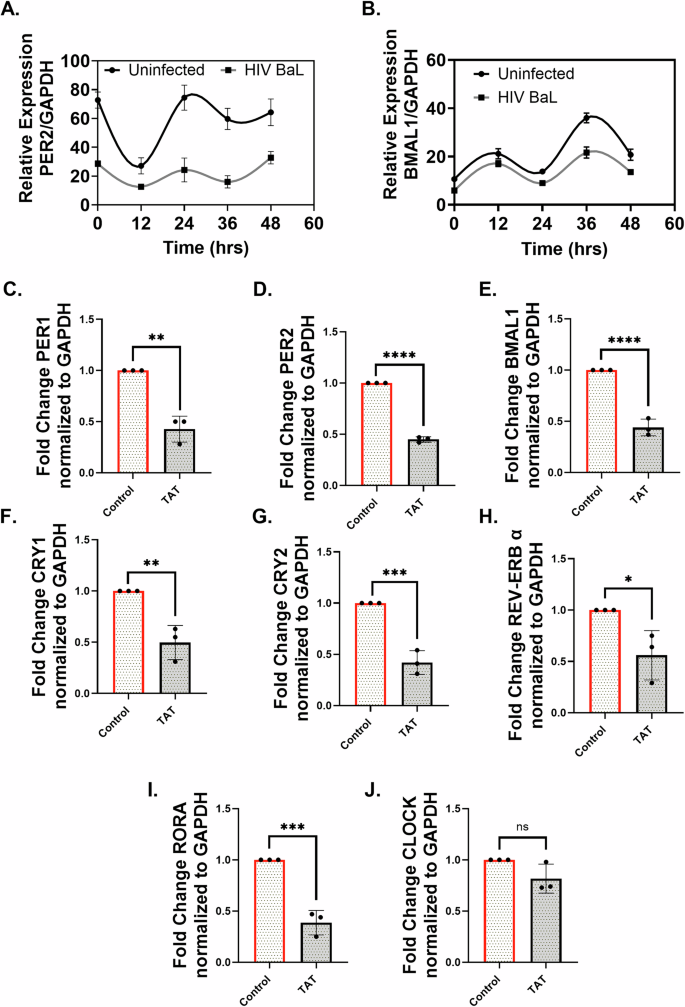
Previous studies have shown that levels of HIV TAT in the cerebrospinal fluid of PLWH negatively correlate with circadian rhythms, affecting sleep as well as blood pressure22, and that TAT expression in the brain alters circadian regularity and activity23. Our findings, along with previous reports, have demonstrated that the bronchial epithelium expresses canonical HIV entry receptors and is permissive to HIV infection24,25. HIV infection of NHBE ALI cultures demonstrated suppression of BMAL1 and PER2 while also showing a weak amplitude of circadian oscillations compared to the control cultures (Fig. 2A, B).
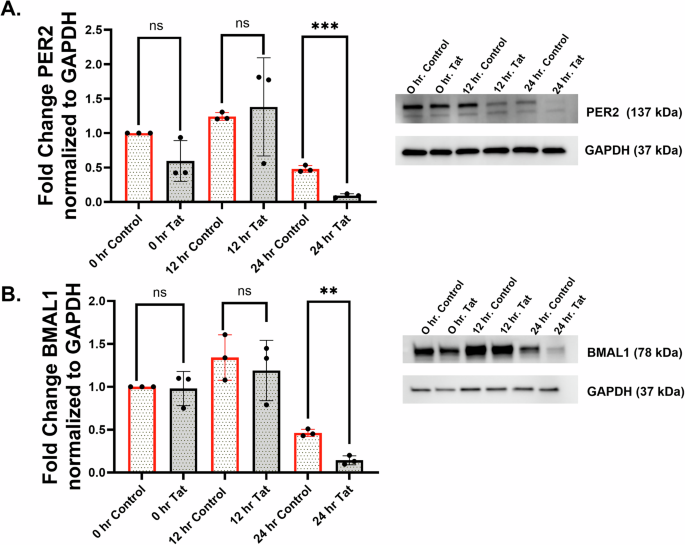
To determine any specific effects of TAT in disrupting the circadian rhythms of airway epithelia, we treated NHBE ALI cells with recombinant HIV TAT and quantified the expression of clock genes. All the core clock genes (PER1, PER2, BMAL1, CRY1, CRY2, REV-ERBα, and RORΑ) showed significant suppression following TAT treatment compared to the control (Fig. 2C–I). Although the suppression of CLOCK trended lower with TAT treatment, it was not significant (Fig. 2J). To understand the temporal effects of HIV TAT on the circadian expression pattern of BMAL1 and PER2 at the protein level, the circadian clock was synchronized following TAT treatment. The expression of the BMAL1 and PER2 proteins in the TAT-treated group demonstrated a significant decrease compared to the heat-inactivated TAT controls (Fig. 3A, B). Both BMAL1 and PER2 proteins showed higher expression at the 12 h time point and lower expression at the 24 h time point. Lower protein expression was observed with TAT treatment at 0, 12, and 24 h for PER2, while maximum downregulation of BMAL1 was observed at 24 h.
HIV TAT suppresses SIRT1 in NHBE ALI cells
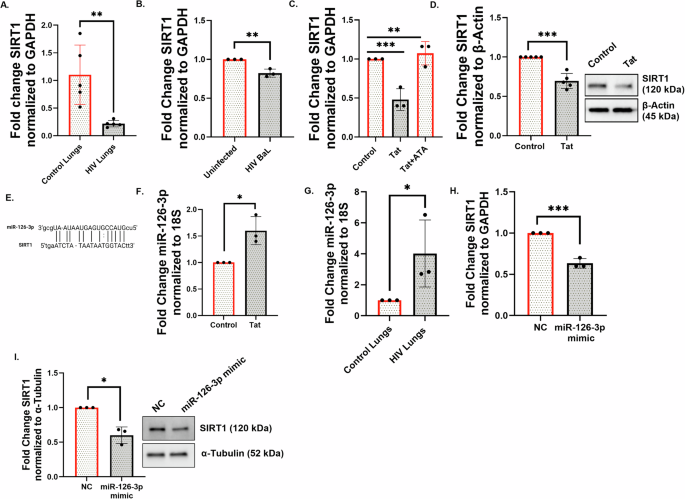
SIRT1 is a master clock deacetylase26. We have previously demonstrated that disruption of SIRT1-mediated BMAL1 regulation in the epithelium plays an important role in cigarette smoking-induced lung inflammation18. SIRT1 is a highly conserved class III histone deacetylase that is involved in deacetylation of several proteins27. Kwon H-S et al. (2009) showed that TAT promotes hyperactivation of T-cells by inhibiting the SIRT1 activity28. While this interaction was at the protein level, we wanted to explore whether other mechanisms exist by which HIV dysregulates SIRT1. We previously reported that HIV TAT dysregulates the bronchial epithelial microRNAome29. We explored whether HIV TAT and, by extension, HIV infection, suppresses SIRT1 biogenesis in bronchial epithelial cells and if this is due to miRNA-mediated silencing. We first investigated if SIRT1 levels are suppressed in PLWH. We quantitated SIRT1 mRNA levels in control lungs and lungs from HIV-positive donors. Figure 4A shows that SIRT1 is suppressed in lungs of HIV-positive donors compared to HIV-negativecontrols.
To further validate these findings, we infected NHBE ALI cultures with HIV and quantitated SIRT1 RNA levels. Figure 4B demonstrates that SIRT1 is downregulated in the HIV-infected NHBE ALI cultures. Our data show that HIV infection suppresses SIRT1 both in vivo in human lung tissue and in vitro in NHBE ALI cultures. NHBE ALI cells were treated with HIV TAT (or control). Another experimental group was pre-treated with aurintricarboxylic acid (ATA), a small molecule inhibitor of the miRNA processing enzyme Drosha30. Total RNA was quantitated for SIRT1 expression 48 h post TAT treatment. The qRT-PCR data confirmed the downregulation of SIRT1 mRNA and its reversal by ATA, suggesting that TAT suppresses SIRT1 by miRNA-mediated regulation (Fig. 4C). Similarly, NHBE ALI cultures treated with HIV TAT demonstrated decreased levels of SIRT1 protein similar to mRNA suppression (Fig. 4D).
HIV TAT upregulates miR-126-3p, suppresses SIRT1, and increases acetylated BMAL1
We previously showed that HIV TAT treatment leads to statistically significant changes in the expression of 83 different microRNAs in bronchial epithelial cells21. We used miRTARbase and miRanda to identify potential miRNA-SIRT1 pairs31. Using miRanda, the miRNA-target prediction confirmed the presence of a complementary sequence (including the seed sequence) at position 922–943 for the miR-126-3p binding site on the 3’UTR of the SIRT1 mRNA (Fig. 4E). This sequence is highly conserved among several vertebrates, suggesting a potential novel miRNA-mRNA interaction for microRNA-mediated SIRT1 regulation. We revisited our earlier miRNA array data and noticed that HIV TAT also upregulated miR-126-3p (log2 fold change of 4.81; p-value of 0.059, narrowly missing statistical significance; supplementary data in ref. 21). We re-examined this finding to determine if miR-126-3p plays a role in HIV TAT-mediated SIRT1 suppression and observed that miR-126-3p levels were significantly increased in TAT-treated NHBE cells (Fig. 4F). Lung tissues from HIV-positive donors also demonstrated significant upregulation of miR-126-3p compared to the controls (Fig. 4G). Finally, we validated the miR-126-3p-SIRT1 regulatory axis by transfecting miR-126-3p miRNA mimics (or control) in BEAS-2B airway epithelial cell lines. Our data show that miR-126-3p transfection suppresses SIRT1, thereby confirming the regulation of SIRT1 by miR-126-3p (Fig. 4H, I). Together, these data identify miR-126-3p as a novel microRNA candidate for HIV TAT-mediated SIRT1 suppression.
Nakahata et al. showed that SIRT1 regulated the activity of the CLOCK-BMAL1 heterodimer by deacetylating BMAL132. We have previously shown that dysregulation of the SIRT1/BMAL1 axis in airway epithelial cells causes lung inflammation in mice exposed to cigarette smoke18. To determine if SIRT1 suppression translates to changes in the acetylation status of BAML1, its deacetylase target, we transfected NHBE cells with siSIRT1. SIRT1 silencing was confirmed by qRT-PCR and western blot analyzes of SIRT1 levels (Supplementary Fig. 2A, B). SIRT1 silencing led to a significant increase in levels of Ac-BMAL at Lys538 compared to the control (Supplementary Fig. 2C). Ac-BMAL1 further manifests as upregulation of the IL-6 gene, as previously reported33,34 (Supplementary Fig. 2D).
HIV TAT disrupts circadian oscillation by upregulating miR-126-3p
To investigate the role of TAT and miR-126-3p on PER2 expression and circadian oscillation in bronchial epithelial cells, we transduced NHBE cells with the lentiviral promoter reporter vector, PER2-Luc, in which the PER promoter drives the firefly luciferase reporter. Undifferentiated bronchial epithelial cells were used since we have previously shown efficient transduction of undifferentiated cells with lentiviral vectors35, but poor transduction of differentiated bronchial epithelial cells (not shown). Firefly luciferase is used extensively across various organisms and cell types to study circadian rhythm36,37,38. To determine the role of miR-126-3p in dysregulating circadian rhythm via SIRT1 suppression, we transfected PER2-transduced NHBE cells with miR-126-3p. Our data show circadian expression of luciferase in respective controls with maximal PER2 promoter activity recorded at 12-h and 36-h time intervals. Treatment with HIV TAT (Supplementary Fig. 3A) or transfection with miR-126-3p mimic (Supplementary Fig. 3B) significantly depressed the amplitude and levels of luciferase expression driven by PER2 promoter expression compared to the controls. SIRT1 levels were quantitated for the peak amplitude time point (36 h) and showed that HIV TAT and miR-126-3p suppress SIRT1. These data demonstrate that HIV TAT induces miR-126-3p to suppress SIRT1 with consequent downstream effects on lung circadian gene expression. We have used undifferentiated as they show increased transfection efficiency compared to fully differentiated NHBE ALI culture. We do not anticipate that the use of differentiated cells would alter the overall results or conclusions, as the mechanism of RNA interference is highly conserved across cell types and species.
TAT-mediated disruption of circadian rhythm leads to increased secretion of proinflammatory cytokines
We have demonstrated that disruption of the SIRT1/BMAL1 axis in the airway can lead to increased inflammation18. NHBE ALI cultures were treated with TAT or control for 48 h as described in the methods. Similarly, another set of cells was transfected with miR-126-3p or control miR-NC. TAT treatment significantly increased proinflammatory cytokines in NHBE ALI cultures compared to the control (Supplementary Fig. 4A). Specifically, we observed increased levels of TNF-α, IL-6, IL-8, GM-CSF, bFGF, IFN-γ, IL-4, and IL-17 in the TAT-treated cells compared to controls. Transfection with miR-126-3p mimics upregulated only MCP-1 and IFN-γ (Supplementary Fig. 4B; both cytokines were also upregulated by TAT) compared to multiple cytokines upregulated by HIV TAT. Separately, the fold change in CXCL5 mRNA expression was higher in TAT-treated NHBE cells compared to control cells as determined by qRT-PCR (Supplementary Fig. 4C). These cytokines play a significant role in lung inflammation, specifically in patients with COPD39,40,41,42,43,44,45.
SIRT1 knockout disrupts circadian protein expression and enhances HIV-1-induced inflammation
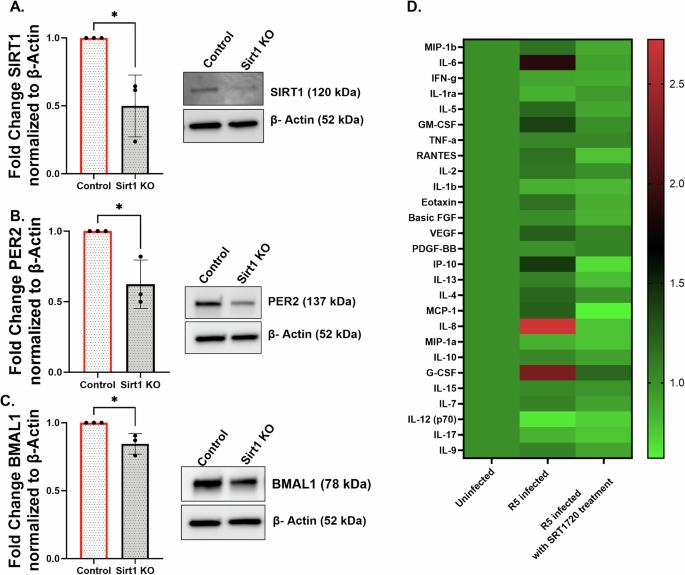
To investigate the role of SIRT1 in regulating circadian clock proteins and inflammation, we generated SIRT1 knockout NHBE cultures using CRISPR/Cas9 and confirmed the knockout via western blot analysis. SIRT1 protein levels were significantly reduced in the knocked-out cells compared to the control group (Fig. 5A); however, a low level of residual SIRT1 expression was still detected, indicating incomplete knockout. Next, we examined the SIRT1 depletion on core circadian components. Western blot analysis revealed a significant reduction in PER2 (Fig. 5B) and BMAL1 (Fig. 5C) protein levels upon SIRT1 knockout, indicating that SIRT1 is essential for maintaining circadian rhythm stability in NHBE ALI cultures and directly regulates the core clock genes PER2 and BMAL1.
Additionally, we assessed the inflammatory response in NHBE cultures in different conditions using Luminex analysis. HIV-1 (R5) infection led to a significant increase in proinflammatory cytokine secretion compared to the control group (Fig. 5D), demonstrating a virus-induced inflammatory response. However, pre-treatment with 1 μM SRT1720 (R5+SRT1720), a potent SIRT1 activator, resulted in a marked reduction in cytokine levels compared to the R5-infected group. Analysis of proinflammatory cytokines from the culture supernatant demonstrated that HIV infection increased expression of IL-6, IL-8, G-CSF, GM-CSF, MCP-1, and IP-10. Treatment with SRT1720 reversed these increases in proinflammatory cytokines. In addition, we also observed a marked reduction in the baseline levels of several key proinflammatory cytokines, including IL-5, RANTES, VEGF, and IL-4 (Fig. 5B).
Circadian gene suppression by HIV TAT leads to lung inflammation and features of emphysema in SP-C TAT mice
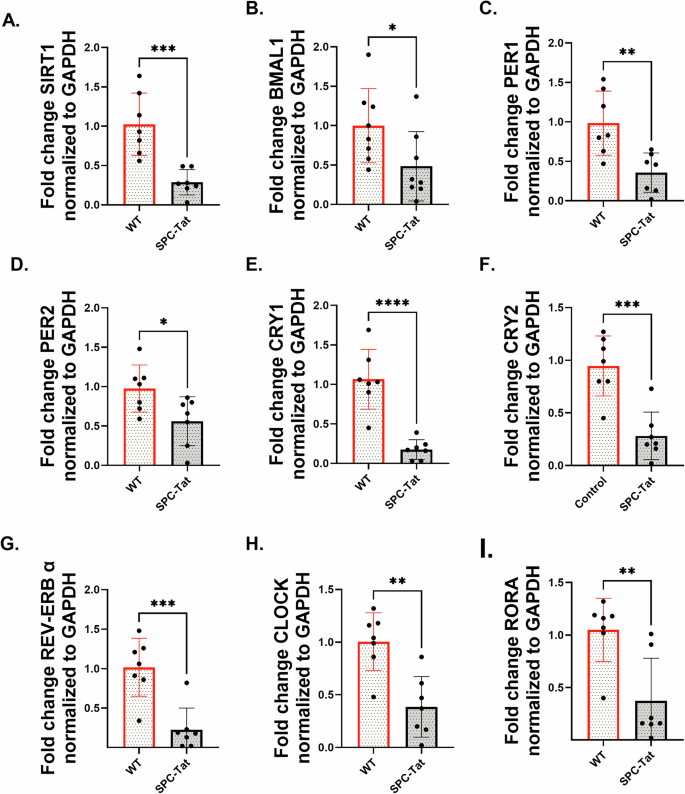
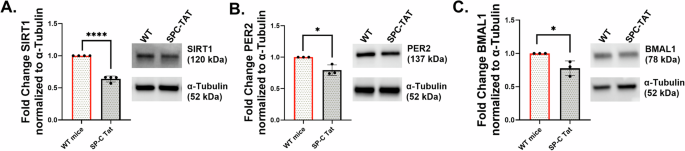
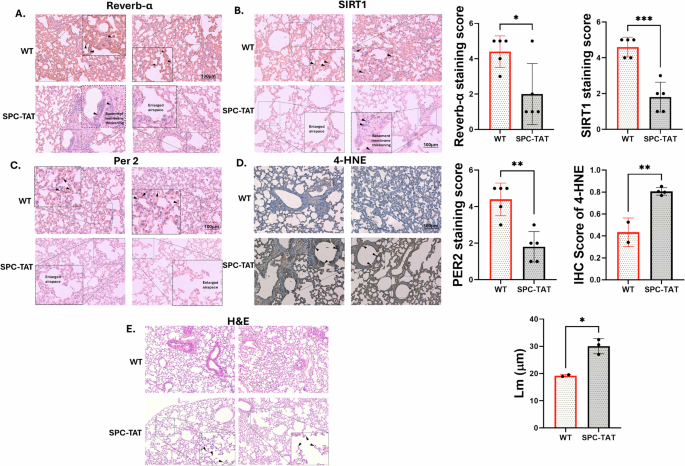
SP-C mice were first generated and described by Cota-Gomez et al.46, with TAT (86-amino acid isoform) expressed under the control of surfactant protein-C promoter (SP-C), thereby restricting TAT expression to the lung and, more specifically, the small airways. HIV TAT contains a protein transduction domain that allows it to be secreted and taken up by other cells47,48,49,50, and can have pleiotropic effects on other lung cells as well. PLWH demonstrates the onset of COPD symptoms as early as age 38-4751,52. Lungs from 7-month-old SP-C TAT mice (~35 years human equivalent53) were analyzed for expression of major circadian genes and SIRT1. SP-C TAT mice demonstrated a significant downregulation of core circadian genes SIRT1, BMAL1, PER1, PER2, CRY1, CRY2, REV-ERB α, CLOCK, and RORA normalized with GAPDH (Fig. 6A–G). These results are similar to those observed by us in lungs from HIV-positive lung donors (Fig. 1A–H), confirming the role of HIV TAT in circadian disruption in the lung. Western blot analyzes demonstrated suppression of SIRT1 protein levels (Fig. 7A) and its circadian deacetylase targets BMAL1 and PER2 (Fig. 7B, C) in lungs from SP-C TAT mice compared to WT littermate controls. IHC analysis of SPC-TAT mice was carried out to determine the impact of HIV-related stress on tissue architecture and molecular clock protein expression under in vivo conditions. Lungs from control and SP-C TAT-expressing mice were examined using specific antibodies against molecular clock proteins. Histological evaluation of airway/alveolar and parenchymal lung sections revealed significant downregulation of molecular clock proteins (PER2, SIRT1, and REV-ERBα), consistent with mRNA expression data (Fig. 8A–C). Surprisingly, BMAL1 does not show any significant downregulation in IHC staining (data not shown). Protein expression was localized primarily in the epithelial and basement membrane regions, suggesting that these clock proteins regulate essential cellular functions in these areas. Moreover, we also observed abnormal enlargement of the alveolar airspaces and epithelial hyperplasia in the airway, with the cells surrounding the airway regions exhibiting prominent thickening. Lungs from SP-C TAT mice demonstrated increased ROS damage and airspace enlargement (evident from the mean linear intercept) compared to WT littermate controls, characteristic of early stages of emphysema (Fig. 8D, E). These data demonstrate that HIV TAT-mediated circadian dysregulation leads to lung inflammation and features of emphysema.
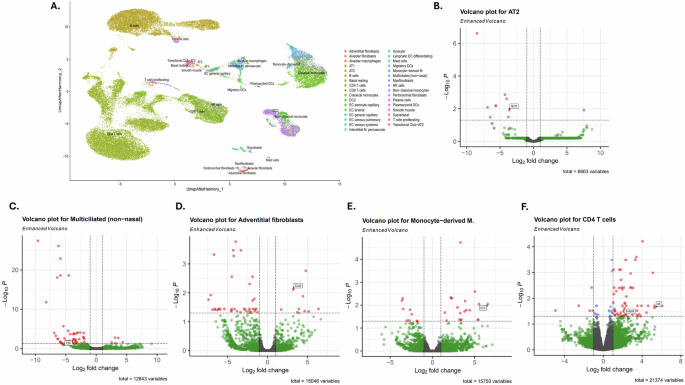
Single-cell RNA sequencing of SPC-TAT mouse lungs
To investigate early events, pathways, and cell types involved in HIV TAT-associated lung inflammation as a function of transcriptomic changes, single-cell RNA sequencing (ScRNAseq) was performed on lungs from 4-month-old (juvenile) SP-C TAT mice and age-matched wild-type controls. Following quality control and filtering, we obtained high-quality single cells across all samples. Cell clustering and annotation revealed the major cell types: AT2 cells, multiciliated cells, adventitial fibroblasts, endothelial cells, macrophages, and T-cells. To account for batch effects, sample integration was performed using the Harmony algorithm, resulting in a unified UMAP representation. Figure 9A is a UMAP visualization identifying major immune, epithelial, endothelial, and stromal populations. Integration was performed on a merged Seurat object that had undergone normalization, identification of highly variable features, and principal component analysis. Key annotated clusters include AT2, AT1, multiciliated cells, alveolar macrophages, fibroblast subtypes, and diverse immune subsets. We observed SIRT1 suppression in AT2 cells in SP-C Tat mice (adjusted p-value of 0.012) (Fig. 9B). In line with our earlier data on suppression of CFTR by HIV TAT, we observed CFTR suppression in ciliated epithelial cells (adjusted p-value of 0.004) of SP-C TAT mice24,29 (Fig. 9C). Ciliated cells in the airway epithelium also demonstrated a non-significant reduction of core clock genes including REV-ERBα (NR1D1), BMAL1, PER1, CRY1, and RORA (Fig. 9C). Surprisingly, SIRT1 and PER2 levels were mildly elevated in ciliated cells (not significant), suggesting a possible compensatory mechanism in the early stages of these mice. Given that NR1D1 regulates metabolism and SIRT1 expression and activity are regulated by metabolism, decreased NR1D1 could lead to the small compensatory increase in SIRT1 levels.we did not observe secretion of proinflammatory cytokines in airway epithelial cells. Elevated expression of chemokines and proinflammatory cytokines was observed in adventitial fibroblasts (Fig. 9D), suggesting that lung fibroblasts may play an early role in inflammation in PLWH. Specifically, we found a considerable upregulation of CCL2 (MCP-1) in adventitial fibroblasts (adjusted p-values of 0.007). We also observed increased inflammation in CD4 T-cells, with a significant upregulation of IL5—in SP-C TAT mice (adjusted p-values of 0.0023) (Fig. 9E). The monocyte derived macrophages shows upregulation of IL13 (Fig. 9F). These data suggest that pulmonary fibroblasts, T-cells and macrophages may play an important initial role in lung inflammation in PLWH.
Discussion
We and others have shown that circadian dysregulation in the lung can manifest as chronic lung inflammation18,54,55. Chromic lung inflammation is a hallmark of COPD56. COPD is one of the principal comorbidities in PLWH, with increased exacerbations and worse outcomes in HIV-positive smokers. Several studies have confirmed that the prevalence of COPD is higher in PLWH. PLWH often experience disturbed sleep and motor-cognitive issues due to circadian rhythm disorder57. HIV TAT disrupts the circadian system, affecting body temperature, hormone secretion, and immune cell circulation. Studies show that HIV TAT in the brain alters the sleep cycle. However, the molecular mechanisms underlying this effect, specifically the direct effects of HIV proteins on peripheral clocks in organs that have the potential to promote inflammation, have yet to be determined.
Our studies with NHBE cells, HIV-positive donor lungs, and SP-C TAT mice demonstrate for the first time that HIV infection dysregulates the molecular components of the lung circadian clock by an aberrant TAT-induced microRNAome. This microRNAome exhibits decreased SIRT1 levels, with consequent effects on the lung molecular clock genes. The circadian clock in the lungs, as well as the airway epithelium, plays an important role in maintaining inflammation homeostasis. We have demonstrated that disruption of this epithelial circadian clock is associated with lung inflammation and COPD18. To our knowledge, this is the first report that demonstrates the molecular mechanisms underlying HIV TAT-mediated lung inflammation, dysregulation of the lung circadian clock, and COPD development in PLWH.
Previous studies, including ours, have demonstrated that SIRT1 is the master circadian deacetylase and controls acetylation status of circadian genes, consequently affecting their activity as well as expression of the downstream molecular signaling cascade18,58,59. Given previous reports of HIV (and TAT) dysregulating the central clock and affecting sleep patterns, we first examined lungs from PLWH donors for changes in the expression of circadian genes and SIRT1 deacetylase. Surprisingly, we observed suppressed RNA levels of almost all core circadian genes as well as SIRT1. Next, we examined whether HIV infection dysregulates circadian oscillations of BMAL1 and PER2, two clock genes that are targets of SIRT1 deacetylase activity. Infection of NHBE ALI cells with HIV in culture demonstrated a decrease in levels of clock genes and the amplitude of circadian oscillations. Recombinant HIV TAT likewise demonstrated suppressed RNA levels of all circadian genes, and oscillation as well as protein levels of the SIRT1 targets BMAL1 and PER2. While previous reports have shown that HIV TAT can inhibit SIRT1 activity28,60, none of the reports showed that HIV TAT can also suppress SIRT1 biogenesis. Given that SIRT1 is a master deacetylase of the circadian genes and that SIRT1 suppression is associated with suppression of other clock genes leading to lung inflammation18, we examined the mechanism by which HIV TAT suppresses SIRT1. Our data demonstrated that HIV TAT-mediated SIRT1 suppression could be rescued by blocking microRNA processing. We previously reported that HIV TAT dysregulates the bronchial epithelial microRNAome29. We used miRNA-target algorithms to identify miR-126-3p as a potential regulator of SIRT1. In our earlier screen of the aberrant microRNAome by Dutta et al.29, this microRNA upregulation was borderline not significant (p = 0.0595). Using specific probes, we validated the statistically significant upregulation of this microRNA in NHBE ALI cultures treated with recombinant TAT as well as in lungs from HIV-positive donors. The miRNA-target relationship for miR-126-3p and SIRT1 was validated by demonstrating SIRT1 suppression in BEAS-2B airway epithelial cells transfected with miR-126-3p mimic. In our earlier studies, we also demonstrated that HIV upregulates miR-34a-5p as a microRNA that regulates SIRT161. In this study, we show that miR-126-3p, another microRNA upregulated by HIV TAT, also suppresses SIRT1. To the best of our knowledge, this is the first report that demonstrates miR-126-3p-mediated SIRT1 regulation. Our data showing that miR-126-3p alone can alter circadian gene expression and induce proinflammatory cytokines suggest that the actions of miR-34a-5p and miR-126-3p can regulate SIRT1 independently of each other. Our data also show that miR-126-3p mimics demonstrate a greater degree of SIRT1 suppression compared to miR-34a-5p mimic. The experiments required to disentangle the cooperative or independent effects of miR-34a-5p and miR-126a-3p are beyond the scope of this manuscript.
We used multiple experimental approaches to establish the relationship between TAT-mediated SIRT1 suppression and circadian dysregulation. siRNA-mediated SIRT1 suppression led to a corresponding increase in acetylated-BMAL1, and PER2-luciferase reporter assays demonstrated that TAT, as well as miR-126-3p, decreased the levels and amplitude of luciferase reporter oscillations. HIV TAT-mediated SIRT1 suppression and miR-126-3p mimic transfection led to increased secretion of proinflammatory cytokines. While we observed more cytokines upregulated by HIV TAT than miR-126-6p, this may be due to the model system used (NHBE for TAT versus BEAS-2B cells for miR-126-3p mimic). Alternatively, it is possible that TAT has dual effects on SIRT1 in that it suppresses SIRT1 biogenesis (mRNA levels) as well as activity. In contrast, miR-126-3p suppresses only SIRT1 biogenesis, leading to a more robust downregulation of SIRT1 effects with TAT than with miR-126-3p. Of note, MCP-1 (CCL2) and IFN-γ were induced by both TAT and miR-126-3p. We observed that treatment with SRT1720 completely reversed expression of the proinflammatory cytokines IL-6, GM-CSF, IP-10, MCP-1, IL-8, and G-CSF induced by HIV infection. We also observed a general decrease in baseline levels of several key proinflammatory cytokines.
Finally, we validated the effects of HIV TAT in 7-month-old SP-C TAT transgenic mice in which TAT expression is restricted to the airways. We observed an identical pattern of suppression in the circadian genes in lungs from SP-C TAT transgenic mice to that we observed with HIV-positive human donor lungs, with statistically significant suppression of BMAL1, CRY1, CRY2, PER1, PER2, REV-ERBα (NR1D1), CLOCK, RORA and SIRT1. Histology experiments with paired lungs showed that SP-C TAT mice demonstrated decreased SIRT1, PER2, and REV-ERB α expression, and increased oxidative burden (4HNE staining; Fig. 8D) and airspace enlargement characteristic of emphysema. Our data demonstrate that HIV TAT, and by extension, HIV infection, induces miR-126-3p to suppress SIRT1. SIRT1 suppression then dysregulates the core lung circadian clock which then manifests as suppression of other clock genes, including REV-ERBα, leading to inflammation and features of emphysema. While the exact mechanism by which clock dysregulation leads to inflammation remains to be determined, studies have shown that REV-ERBα suppression leads to inflammation in multiple tissues. Another mechanism may be a direct effect of BMAL1 suppression. The CLOCK gene is a positive regulator of, and associates with NFκβ62. SIRT1-mediated BMAL1 deacetylation counteracts CLOCK-dependent NF-κβ activation62. Alternatively, SIRT1 can directly deacetylate p65 to inhibit NF-κβ63. Hence, a combination of SIRT1 suppression and circadian dysregulation can lead to increased NF-κβ activity and decreased REV-ERBα levels, resulting in lung inflammation.
COPD is a multifactorial disease, with multiple interconnected pathways involved in lung inflammation and COPD development. Our data demonstrate that SIRT1 suppression and circadian dysregulation in the lung play an important role in the inflammatory process. PLWH present with emphysema as early as 38–47 years51,52. Understanding early events and cell types involved in lung inflammation can help arrest the development of emphysema. We focused on early transcriptomic changes that may point to initial pathways and cell types involved in lung inflammation, carrying out scRNAseq experiments on ~4-month-old lungs (human age equivalent ~20–25-year-old53). The scRNAseq experiment confirmed the suppression of SIRT1 in AT2 cells in 4-month old mice. ScRNAseq data also validated the HIV TAT-mediated CFTR suppression in epithelial cells previously reported by us29, and identified that this suppression occurs in ciliated cells of the airway epithelium (Fig. 9C). We also observed suppression of clock genes, including REV-ERBα (NR1D1), in ciliated cells. However, at this early age, the suppression had not reached statistical significance yet (not shown). We saw several other cell types driving inflammation in SPC-Tat mice. We observed elevated expression of chemokines and proinflammatory cytokines in adventitial fibroblasts, suggesting that ciliated cells and lung fibroblasts may be early drivers of lung inflammation in PLWH. Of note, we found elevated CCL2 (MCP-1) expression in adventitial fibroblasts in TAT mice. CCL2 is increased in emphysema and is produced in response to inflammatory stimuli by a variety of cells64. Higher CCL2 induces an inflammatory cytokine response and pulmonary leukocyte accumulation. CCL2 increase is also associated with other lung diseases such as pulmonary hypertension, acute lung injury, and pulmonary fibrosis65,66. CCL2 is known to activate CCR2 on monocytes and T cells67. T-lymphocytes are the predominant inflammatory cells present in the central airways and lung parenchyma of COPD patients, and their presence correlates with the degree of alveolar damage and the severity of airway obstruction in these individuals68. CD4+ T cells show significant upregulation of IL5. IL5 plays an important role in driving airway eosinophilic inflammation in both asthma and COPD69. Monocyte-derived macrophages demonstrate upregulation of IL13 which promotes mucus cell hyperplasia70, and airway remodeling by driving fibroblast proliferation71. Identifying these multiple pathways leading to COPD pathology and their links to lung circadian dysregulation is beyond the scope of this manuscript.
A recent concise review by Shafaati et al.72 examined the interplay between HIV infection or HIV-encoded proteins and host circadian timing systems, including molecular clock regulators such as CLOCK. BMAL1, PER/CRY, and REV-ERBα, and discussed how HIV may perturb sleep-wake cycles and antioxidant defenses72. However, our data identifies a mechanism by which HIV dysregulate peripheral clocks (in our case, lungs) leading to inflammation. We further believe that a similar mechanism may underlie dysregulation of the central SCN clock, thereby linking our findings to the broader systemic effects proposed in that work. In conclusion, our data demonstrate for the first time that HIV TAT dysregulates the lung molecular clock to promote inflammation. Further understanding of the downstream pathways can help identify molecular targets to mitigate lung inflammation. Resetting the lung molecular clock using REV-ERBα and SIRT1 activators can serve as therapeutic leads to mitigate lung inflammation, thereby arresting the development and progression of emphysema.
Methods
Sex as a biological variable
Our study did not include sex as a biological variable. Data from both male and female humans and transgenic mice models were analyzed to reach our conclusion. It is important to note that similar trends were observed in both sexes.
Ethics statement
The current study received approval for obtaining de-identified human lung tissue samples through the Material Transfer Agreement and Procurement (Institutional Review Board, IRB) and complied with laboratory protocol set by the Institutional Biosafety Committee (IBC) at the University of Rochester Medical Center. The study (ID: STUDY00006571) was approved on 5 October 2021 by the IRB at the University of Rochester, NY, and is exempted from NIH, DHHS, and FDA regulations. No study subjects or patients were directly involved in this work, as the lungs were procured from third-party sources, as mentioned below. All subjects/patients were 21 years of age or older. All research was conducted in accordance with the applicable guidelines and regulations of the University of Rochester, Rochester, NY.
Human lung tissue
Human lung tissue samples from both non-diseased and diseased (HIV-positive) individuals were obtained from two cohorts - (i) National Disease Research Interchange (NDRI) following the procedures outlined in our previous studies73,74 and (ii) United Network for Organ Sharing facilitated by the International Institute for the Advancement of Medicine (IIAM), which provides fresh non-transplantable human organs/tissues for research purposes. The clinical characteristics of the subjects used in the current study are shown in Supplementary Table 1. In brief, age- (between 45 and 60 years) specimens from non-diseased and diseased individuals with no history of smoking were included in this study. As the ethnicities of HIV-positive samples varied, we excluded these as confounders. None of these donors had a history of alcohol use. The ART (antiretroviral therapy) status of the lung donor is unknown.
Cell cultures and cell lines
NHBE cells were obtained from the University of Miami Life Alliance Organ Recovery Association (LAORA) and Lonza Bioscience (OR, USA). All experiments were conducted using NHBE cells. These primary cultures undergo mucociliary differentiation at the air-liquid interface (ALI), mimicking the in vivo structure and essential physiological functions to regenerate the native bronchial epithelium in an ex vivo setting. NHBE cells were sourced from the lungs of non-smoker donors who tested negative for HIV, Epstein-Barr virus (EBV), cytomegalovirus (CMV), and hepatitis B (HBV) to minimize potential confounding variables. NHBE cells were cultured on collagen I-coated tissue culture dishes in bronchial epithelial growth medium (BEGM). For NHBE ALI cultures, 350,000 NHBE cells per well were differentiated 7–10 days after seeding using collagen IV-coated 12 mm transwell or snapwell clear permeable supports within an ALI culture system. The ALI medium was prepared as previously described75,76. Bronchial epithelial cells from paired lungs were used for control and treatments for all experiments. The bronchial epithelial cell line BEAS-2B was sourced from the American Type Culture Collection (ATCC, USA) and maintained in BEGM basal medium according to the manufacturer’s instructions.
Virus strains and protein
The R5-tropic HIV BaL strain (CCR5 receptor-utilizing HIV) was used throughout the study. NHBE ALI cultures grown on transwells were infected with the R5 tropic HIV strain (BaL) (5 ng/ml p24-equivalent) both apically and basolaterally, as described previously24. HIV-1 IIIB TAT recombinant protein (BEI Resources, NIH HIV Reagent Program, cat. no. HRP-2222) was obtained from the NIH AIDS Reagent Program and reconstituted in PBS with 1 mg/ml BSA and 0.1 mM DTT to achieve the stock concentration of 10 µM.
Circadian expression measurement of genes in NHBE ALI culture
NHBE ALI cultures were either infected with HIV or treated with HIV-1 TAT recombinant protein for the circadian measurement of genes. To synchronize the circadian clock, cells were treated with 10 µM forskolin on both the apical and basolateral surface for 30 min, 24 h after initial treatment. Forskolin induces a transient rise in intracellular cAMP, which resets the molecular clock in peripheral cells77. Following forskolin-based synchronization, cells were washed, and experiments were terminated at designated time points.
microRNA mimic and siRNA transfection
hsa-miR-126-3p mimic (mirVana miRNA mimic, sequence- UCGUACCGUGAGUAAUAAUGCG, cat. no. AM17000), mirVana miRNA mimic negative control (cat. no. 4464058), and siSIRT1 (cat. no. AM16708) were purchased from Thermo Fisher Scientific, MA, USA. BEAS-2B or NHBE cells were seeded on collagen-coated 12-well tissue culture plates (Fisher Scientific). BEAS-2B cells were electroporated with 40 nM miR-126-3p and miR-NC using the Neon Transfection System (Invitrogen, MA, USA), following the manufacturer’s protocol. In another experimental set, NHBE cells were transfected with 40 nM siSIRT1 and NC using lipofectamine RNAiMAX with Opti-MEM medium. Total RNA and protein were collected 48 h post electroporation for quantitative RT-PCR and western blot analysis, respectively.
CRISPR knockout of SIRT1 in NHBE ALI culture
NHBE cells were cultured at the ALI following standard protocol to achieve differentiation. To knock out SIRT1, we utilized the Dharmacon predesigned SIRT1-specific Edit-R CRISPR guide RNA. NHBE cultures were transfected with the CRISPR guide RNA along with Edit-R Cas9 protein NLS using DharmaFECT Duo Transfection Reagent (cat. no. T-2010-01) following the manufacturer’s instructions. Cells were harvested 48 h post transfection, and total protein was extracted using RIPA buffer supplemented with protease inhibitor.
Quantification of miRNA and mRNA expression by quantitative reverse transcription-PCR (qRT-PCR)
Total RNA was extracted from cells using the PureLink RNA Mini Kit (Invitrogen, cat. no. 12183020). RNA purity and concentration were assessed with a microspot RNA reader (Synergy HT Multi-Mode Microplate Reader, BioTek, USA), ensuring an absorbance ratio of 260/280 nm between 1.8 and 2.0. For cDNA synthesis, 0.5 µg of total RNA was reverse transcribed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, cat. no. 4368814, USA) following the manufacturer’s instructions. Relative mRNA expression was quantified with TaqMan Fast Advanced Master Mix (Applied Biosystems, cat. no. 4444557) in a 20 µL real-time PCR reaction using gene-specific TaqMan primers on the Bio-Rad CFX96 real-time system. The data were normalized to GAPDH (in case of mRNA quantification) or 18S RNA (in case of non-coding RNA; microRNA quantification) and calculated as mean or relative fold change of target gene expression using the comparative CT method.
Plasmid, lentivirus, and transduction and luciferase reporter assays
The human PER2-luciferase reporter contrast (cat. no. HPRM50381-LvPL01) was purchased from Genecopoeia, USA, and packaged as VSV-G pseudo typed lentiviral vectors by Vector Builder (USA). The luciferase reporter is driven by the PER2 promoter. NHBE cells were transduced with the PER2 promoter-reporter lentiviral vector in the presence of polybrene (5 µg/ml). Transduced cells were incubated for 24 h at 37 °C, after which cells were subjected to antibiotic selection using 1 µg/mL puromycin for 72 h to ensure stable integration of the reporter construct.
PER2
Luc-transduced cells were treated with HIV TAT (25 nM) (or control) or transfected with 40 nM miR-126-3p mimic or mirVana miRNA mimic negative control. To synchronize the circadian clock, the cells were treated with 10 µM forskolin for 30 min, as reported by Ramsey et al.77. Following synchronization, the medium was replaced with fresh growth medium, and luciferase activity was measured every 12 h for 48 h using the Dual-Luciferase Reporter Assay system (Promega, cat. no. E2920, USA). The firefly luciferase activity was measured using Synergy HT Multi-Mode Microplate Reader (BioTek).
Western blotting
Proteins were isolated from infected or treated cells using RIPA buffer (Thermo Fisher Scientific, cat. no. #89901) mixed with protease inhibitor cocktail (Thermo Fisher Scientific, cat. no. 78429). Protein concentration was quantified by a nanodrop spectrometer using the BSA method (Thermo Fisher Scientific). Equal amounts of total protein were loaded on 4–20% precast polyacrylamide gels (Bio-Rad, cat. no. 4568094) and run at 100 V, followed by transfer onto a polyvinylidene difluoride membrane. The membrane was blocked with a 10% blocking solution for 1 h. and then incubated with primary antibodies, either SIRT1 (1:1000 dilution; Cell Signaling, cat. no. 9475S), PER2 (1:2500 dilution; Thermo Fisher Scientific, cat. no. MA5-44772/Novus Biologicals, cat. no. NB100-125), BMAL1 (1:1000 dilution; Cell Signaling, cat. no. 14020), anti-BMAL1 (acetyl-lys538) antibody (1:2500 dilution, Antibodies.com, cat. no. A51697), β-actin (1:1000 dilution; Cell Signaling, cat. no. 4970S), α-tubulin (1:2500 dilution; Cell Signaling, cat. no. 2125S), or GAPDH (1:2500 dilution; Cell Signaling, cat. no. 2118S), in 5% blocking solution overnight. Membranes were washed with TBS (Tris-buffered saline, Bio-Rad, cat. no. 1706435) mixed with Tween 20 (Sigma-Aldrich, cat. no. 9005645) and incubated for 1 h with horseradish peroxidase-conjugated anti-rabbit (Promega, cat. no. W4011) or anti-mouse (Promega, cat. no. W4021) secondary antibody (1:2500 dilution) mixed with 1% blocking solution. As described by the manufacturer, the protein bands were detected in Chemidoc (Bio-Rad Laboratories, USA) using the supersignal west femto maximum sensitivity substrate (Thermo Fisher Scientific, cat. no. 34095). The density values were calculated using the Quantity One software (Bio-Rad Laboratories) and normalized to either β-actin, α-tubulin, or GAPDH.
Luminex assay of cell supernatant
The levels of several proinflammatory mediators were assessed using the Luminex multiplex assay. We used the Bio-Plex Pro Human Cytokine 27-plex Assay Kit (BioRad, cat. no. 171AK99MR2) according to the manufacturer’s instructions to estimate different cytokine levels.
SPC TAT mice
The study adhered to all applicable animal care regulations as outlined in the most recent edition of the National Institute of Health’s “Guide for the Care and Use of Laboratory Animals”. Additionally, all procedures were conducted in accordance with approved protocols from the Institutional Animal Care and Use Committee at Florida Institute University (FIU). SPC TAT mice were generated previously46 and supplied by Dr. Christopher Evans at the University of Colorado Anschutz Medical Campus, and further breeding was done at the FIU Animal Care Facility. Transgenics were confirmed by genotyping for SPC-TAT by PCR using tail DNA. Wild-type littermates were used as a control.
Immunohistochemistry (IHC)
IHC was performed as previously described78. Briefly, 0.5 µm-thick paraffin-embedded mouse lung tissue sections were used for IHC. Deparaffinization and hydration of the sections were performed by immersing them twice in 100% xylene, followed by gradient dilution of ethanol (100%, 90%, 70%, and 50%), each for 5 min. Antigen retrieval was carried out using citrate buffer (pH 6). Peroxidase activity was blocked with 3% hydrogen peroxide in methanol for 15 min. After permeabilization and blocking, the sections were incubated overnight at 4 °C with the respective primary antibodies: Sirt11 (1:200; cat. no. NBP1-51641), Reverbα (1:100; cat. no. H00009572-M02), and Per2 (1:50; cat. no. 67513-1-Ig). Frozen human lung sections (0.5 µm) were processed in parallel using the same antigen retrieval, blocking, and peroxidase quenching steps, and were incubated overnight with 4 °C with a Tat-specific antibody (1:100; cat no. ab42359). Following primary antibody incubation, the sections were incubated with HRP-conjugated secondary antibody (1:500 dilution) at room temperature for 1 h. The sections were developed using DAB, and counterstaining was performed with hematoxylin. After dehydration in graded ethanol and xylene, the sections were mounted using DPX and visualized under bright-field microscopy. Images were captured at 20× magnification, and 10 random images were scored via ImageJ based on specific protein expression in the alveolar and airway regions.The relative IHC score assessment based on positive staining intensity was performed in a semi-quantitatively and blinded manner 0 = no staining; 1 = weak staining; 2 = moderate staining; and 3 = intense staining as described by us previously78.
Lung morphometry
Mouse lungs were inflated with 1% low-melting-point agarose at a pressure of 25 cm H2O and then fixed with 4% neutral-buffered formalin without lavage. Dehydrated lung tissue was embedded in paraffin and sectioned (4 µM) using a rotary microtome (Microm International GmbH, Walldorf, Germany). Xylene and graded alcohol were used to rehydrate the lung sections before staining them with hematoxylin and eosin (H&E). As a measure of airspace enlargement or emphysema, the mean linear intercept (Lm) of the airspace was calculated using MetaMorph software (Molecular Devices, USA)79,80. Lm was calculated for each sample on the basis of 10 random field, images taken at 10× magnification. Airway and vascular structures were eliminated from the analysis. Lung sections were stained with anti-4-HNE antibody (Abcam, cat. no. ab48506). Slides were then developed with DAB Quanto Chromogen and Substrate (Thermo Fisher Scientific, cat. no. TA-125-QHDX).
Single-cell preparation from fresh mouse lung
Single-cell suspensions were prepared from 4-month-old SPC TAT mice and their WT littermates using a combination of enzymatic digestion and mechanical dissociation with the GentleMACS Dissociators system, following a protocol described previously (Miltenyi Biotec, USA). Following the gentle MACS program, the suspensions were strained using MACS SmartStrainer and centrifuged at 300 × g for 10 min. Following centrifugation, the cells were resuspended in red blood cell (RBC) lysis buffer to remove red blood cells and incubated at 4 °C for 10 min. The mixture was then diluted with cold PBS containing 10% FBS and centrifuged at 800 × g at 4 °C for 10 min. The dead cells in the final pellet were removed using the Dead Cell Removal Kit according to the manufacturer’s instructions (Miltenyi Biotec, cat. no. 130-090-101).
Library preparation, sequencing, and analysis
Single-cell RNA sequencing was performed at the University of Miami Miller School of Medicine. Approximately 10,000 cells from each lung cell suspension were used for the preparation of the library. 10× chromium libraries were prepared according to the manufacturer’s protocol (10x Genomics). The library was sequenced on NovaSeq 6000 (Illumina, USA) at a mean sequence depth of 150,000 read pairs per cell. ScRNAseq data analysis was conducted by Singulomics Corporation (https://singulomics.com/, Bronx, NY). Viable cell suspensions were loaded into the Chromium Controller (10× Genomics, USA) to create gel beads-in-emulsion (GEM), with each GEM capturing a single cell and barcoded oligonucleotides. The GEMs were then transferred to the SimpliAmp 96-well Thermal Cycler (Thermo Fisher Scientific), in which reverse transcription was carried out within each GEM (GEM-RT). The resulting cDNA was amplified and purified using Silane DynaBeads (10X Genomics) and the SPRI select reagent kit (Backman Coulter, USA). Amplified full-length cDNAs from poly-adenylated mRNA were then used to generate a 3′ gene expression library (Chromium Next GEM 3′ Single Cell Reagent Kit, v3.1), following the manufacturer’s instructions (10× Genomics). Amplified cDNAs and the libraries were measured by the Qubit dsDNA HS assay (Thermo Fisher Scientific), and quality was assessed by BioAnalyzer (Agilent Technologies, USA). Libraries were sequenced on a NovaSeq instrument (Illumina), and reads were subsequently processed using 10× Genomics Cell Ranger analytical pipeline (v7.2.0) and mouse mm10 reference with the addition of the TAT transgene (NC_001802.1). After the filtering process, which retained only cells with a mitochondrial content of 15% or less, with a hemoglobin gene content of 25% or less, UMI counts greater than 500, and the number of features (genes) between 200 and 10,000, the analysis included three samples from the TAT condition (totaling 24,617 cells) and four samples from the control condition (totaling 20,407 cells).
Sample integration was performed using Harmony to correct for batch effects81. A merged Seurat object served as input, incorporating normalized data with predefined highly variable features and principal components. To prepare this merged object, each sample was independently normalized using SCTransform v2, and integration features were identified across samples using Seurat. The normalized Seurat objects were then merged, with variable features designated as integration features, and 20 principal components were computed on the merged dataset. Next, a UMAP representation was generated using RunUMAP(reduction = “harmony”, dims = 1:20), based on the Harmony embeddings. Clustering analysis was subsequently performed using Louvain clustering, implemented with FindNeighbors(reduction = “harmony”, dims = 1:20) and FindClusters(resolution = 0.5), using the Seurat R package82.
To identify cell types, we employed the Azimuth application and Seurat’s anchor-based label transfer method82,83, leveraging a single-cell RNA-seq reference dataset comprising 584,944 cells obtained from human lung. This reference was constructed by integrating data from 107 individuals across 14 datasets, including both published and unpublished data from the Human Lung Cell Atlas (HLCA)84. Cell typing analysis was conducted on each sample individually, annotating at the most detailed level possible using the ann_level_finest from the reference. Annotations from individual samples were then used for the integrated sample analysis. Differential expression analysis was performed using a pseudobulk approach, where cells from each sample were aggregated by cell type to generate sample-level expression profiles. The EdgeR likelihood ratio test was used to compare gene expression between conditions for each cell type, leveraging sample-level replicates to improve statistical power while maintaining single-cell resolution85.
Statistical analyses
Unless otherwise stated, data were expressed as mean ± SEM from at least three different NHBE ALI cultures from paired lungs, SPC-TAT mice or their WT littermates, or human lung donors. Statistical analyzes were performed using Prism 8 (GraphPad Software, USA). The data were analyzed statistically using unpaired t-tests for comparisons between two groups or ANOVA followed by Tuker-Kramer’s honestly difference test for multiple group comparisons, as appropriate. The significance threshold was set at p < 0.05.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data supporting the findings of this study are available within the paper and its Supplementery material. Uncropped and unedited blot images are provided as Supplementery Information. Sequencing data is available through the SRA bioproject PRJNA1363216.
References
-
Organization, W. H. World Health Statistics 2022: Monitoring Health for the SDGs. Sustainable Development Goals 7 (2022).
-
Harper, J. et al. Progress note 2024: curing HIV; not in my lifetime or just around the corner? Pathog. Immun. 8, 115 (2023).
Article PubMed Google Scholar
-
Roomaney, R. A., van Wyk, B. & Pillay-van Wyk, V. Aging with HIV: increased risk of HIV comorbidities in older adults. Int. J. Environ. Res. Public Health 19, 2359 (2022).
Article PubMed PubMed Central Google Scholar
-
Guaraldi, G. et al. Premature age-related comorbidities among HIV-infected persons compared with the general population. Clin. Infect. Dis. 53, 1120–1126 (2011).
Article PubMed Google Scholar
-
Byanova, K., Kunisaki, K. M., Vasquez, J. & Huang, L. Chronic obstructive pulmonary disease in HIV. Expert Rev. Respir. Med. 15, 71–87 (2021).
Article PubMed Google Scholar
-
Tsai, C. L., Brenner, B. E. & Camargo, C. A. Jr Circadian-rhythm differences among emergency department patients with chronic obstructive pulmonary disease exacerbation. Chronobiol. Int. 24, 699–713 (2007).
Article PubMed Google Scholar
-
Scichilone, N. et al. Circadian rhythm of COPD symptoms in clinically based phenotypes. Results from the STORICO Italian observational study. BMC Pulm. Med. 19, 1–13 (2019).
Google Scholar
-
Canonica, G. et al. Characterization of circadian COPD symptoms by phenotype: methodology of the STORICO observational study. Eur. J. Intern. Med. 43, 62–68 (2017).
Article PubMed Google Scholar
-
Ayyar, V. S. & Sukumaran, S. Circadian rhythms: influence on physiology, pharmacology, and therapeutic interventions. J. Pharmacokin. Pharmacodyn. 48, 321–338 (2021).
Article Google Scholar
-
Fagiani, F. et al. Molecular regulations of circadian rhythm and implications for physiology and diseases. Signal Transduct. Target. Ther. 7, 41 (2022).
Article PubMed PubMed Central Google Scholar
-
Zhang, R., Lahens, N. F., Ballance, H. I., Hughes, M. E. & Hogenesch, J. B. A circadian gene expression atlas in mammals: implications for biology and medicine. Proc. Natl Acad. Sci. USA 111, 16219–16224 (2014).
Article PubMed PubMed Central Google Scholar
-
Dibner, C., Schibler, U. & Albrecht, U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 72, 517–549 (2010).
Article PubMed Google Scholar
-
Pickel, L. & Sung, H.-K. Feeding rhythms and the circadian regulation of metabolism. Front. Nutr. 7, 39 (2020).
Article PubMed PubMed Central Google Scholar
-
Sundar, I. K., Yao, H., Sellix, M. T. & Rahman, I. Circadian molecular clock in lung pathophysiology. Am. J. Physiol.-Lung Cell. Mol. Physiol. 309, L1056–L1075 (2015).
Article PubMed PubMed Central Google Scholar
-
Bryant, A. J. et al. A wrinkle in time: circadian biology in pulmonary vascular health and disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 322, L84–L101 (2022).
-
Parlak, G. C. et al. The secondary pocket of cryptochrome 2 is important for the regulation of its stability and localization. J. Biol. Chem. 298, 102334 (2022).
-
Cao, X., Yang, Y., Selby, C. P., Liu, Z. & Sancar, A. Molecular mechanism of the repressive phase of the mammalian circadian clock. Proc. Natl Acad. Sci. USA 118, e2021174118 (2021).
Article PubMed Google Scholar
-
Hwang, J. W., Sundar, I. K., Yao, H., Sellix, M. T. & Rahman, I. Circadian clock function is disrupted by environmental tobacco/cigarette smoke, leading to lung inflammation and injury via a SIRT1-BMAL1 pathway. FASEB J. 28, 176–194 (2014).
Article PubMed PubMed Central Google Scholar
-
Conti, V. et al. Sirtuin 1 and aging theory for chronic obstructive pulmonary disease. Anal. Cell. Pathol. 2015, 897327 (2015).
-
Ying, S.-Y., Chang, D. C. & Lin, S.-L. The microRNA (miRNA): overview of the RNA genes that modulate gene function. Mol. Biotechnol. 38, 257–268 (2008).
Article PubMed Google Scholar
-
Dutta, R., Chinnapaiyan, S., Santiago, M., Rahman, I. & Unwalla, H. Gene-specific MicroRNA antagonism protects against HIV Tat and TGF-β-mediated suppression of CFTR mRNA and function. Biomed. Pharmacother. 142, 112090 (2021).
Article PubMed PubMed Central Google Scholar
-
Wang, T. et al. HIV T at protein affects circadian rhythmicity by interfering with the circadian system. HIV Med. 15, 565–570 (2014).
Article PubMed PubMed Central Google Scholar
-
Duncan, M. J. et al. Effects of chronic expression of the HIV-induced protein, transactivator of transcription, on circadian activity rhythms in mice, with or without morphine. Am. J. Physiol. 295, R1680–R1687 (2008).
Google Scholar
-
Chinnapaiyan, S. et al. HIV infects bronchial epithelium and suppresses components of the mucociliary clearance apparatus. PLoS ONE 12, e0169161 (2017).
Article PubMed PubMed Central Google Scholar
-
Devadoss, D. et al. HIV-1 productively infects and integrates in bronchial epithelial cells. Front. Cell. Infect. Microbiol. 10, 612360 (2021).
Article PubMed PubMed Central Google Scholar
-
Belden, W. J. & Dunlap, J. C. SIRT1 is a circadian deacetylase for core clock components. Cell 134, 212–214 (2008).
Article PubMed PubMed Central Google Scholar
-
Liu, T. F. & McCall, C. E. Deacetylation by SIRT1 reprograms inflammation and cancer. Genes Cancer 4, 135–147 (2013).
Article PubMed PubMed Central Google Scholar
-
Kwon, H.-S. et al. Human immunodeficiency virus type 1 Tat protein inhibits the SIRT1 deacetylase and induces T cell hyperactivation. Cell Host Microbe 3, 158–167 (2008).
Article PubMed PubMed Central Google Scholar
-
Dutta, R. K., Chinnapaiyan, S., Santiago, M. J., Rahman, I. & Unwalla, H. J. Gene-specific MicroRNA antagonism protects against HIV Tat and TGF-beta-mediated suppression of CFTR mRNA and function. Biomed. Pharmacother. 142, 112090 (2021).
Article PubMed PubMed Central Google Scholar
-
Dutta, R. K., Chinnapaiyan, S., Rasmussen, L., Raju, S. V. & Unwalla, H. J. A neutralizing aptamer to TGFBR2 and miR-145 antagonism rescue cigarette smoke-and TGF-β-mediated CFTR expression. Mol. Ther. 27, 442–455 (2019).
Article PubMed Google Scholar
-
Huang, H.-Y. et al. miRTarBase 2020: updates to the experimentally validated microRNA–target interaction database. Nucleic Acids Res. 48, D148–D154 (2020).
PubMed Google Scholar
-
Nakahata, Y. et al. The NAD+-dependent deacetylase SIRT1 modulates CLOCK-mediated chromatin remodeling and circadian control. Cell 134, 329–340 (2008).
Article PubMed PubMed Central Google Scholar
-
Tang, L. et al. BMAL1/FOXA2-induced rhythmic fluctuations in IL-6 contribute to nocturnal asthma attacks. Front. Immunol. 13, 947067 (2022).
Article PubMed PubMed Central Google Scholar
-
Early, J. O. et al. Circadian clock protein BMAL1 regulates IL-1β in macrophages via NRF2. Proc. Natl Acad. Sci. USA 115, E8460–E8468 (2018).
Article PubMed PubMed Central Google Scholar
-
Unwalla, H. J., Ivonnet, P., Dennis, J. S., Conner, G. E. & Salathe, M. Transforming growth factor-beta1 and cigarette smoke inhibit the ability of beta2-agonists to enhance epithelial permeability. Am. J. Respir. Cell Mol. Biol. 52, 65–74 (2015).
Article PubMed PubMed Central Google Scholar
-
Hamada, T. et al. In vivo imaging of clock gene expression in multiple tissues of freely moving mice. Nat. Commun. 7, 11705 (2016).
Article PubMed PubMed Central Google Scholar
-
Welsh, D. K., Imaizumi, T. & Kay, S. A. in Methods in Enzymology Vol. 393 269–288 (Elsevier, 2005).
-
Lin, H.-H., Qraitem, M., Lian, Y., Taylor, S. R. & Farkas, M. E. Analyses of BMAL1 and PER2 oscillations in a model of breast cancer progression reveal changes with malignancy. Integr. Cancer Ther. 18, 1534735419836494 (2019).
Article PubMed PubMed Central Google Scholar
-
Pervakova, M. Y. et al. High serum level of IL-17 in patients with chronic obstructive pulmonary disease and the alpha-1 antitrypsin PiZ allele. Pulm. Med. 2020, 9738032 (2020).
-
Aaron, S. D. et al. Granulocyte inflammatory markers and airway infection during acute exacerbation of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 163, 349–355 (2001).
Article PubMed Google Scholar
-
Hussein, F. G., Mohammed, R. S., Khattab, R. A. & Al-Sharawy, L. A. Serum interleukin-6 in chronic obstructive pulmonary disease patients and its relation to severity and acute exacerbation. Egypt. J. Bronchol. 16, 1–11 (2022).
Article Google Scholar
-
Tan, Y. et al. FGF2, an immunomodulatory factor in asthma and chronic obstructive pulmonary disease (COPD). Front. Cell Dev. Biol. 8, 223 (2020).
Article PubMed PubMed Central Google Scholar
-
Southworth, T. et al. IFN-γ synergistically enhances LPS signalling in alveolar macrophages from COPD patients and controls by corticosteroid-resistant STAT1 activation. Br. J. Pharmacol. 166, 2070–2083 (2012).
Article PubMed PubMed Central Google Scholar
-
Huang, A.-X., Lu, L.-W., Liu, W.-J. & Huang, M. Plasma inflammatory cytokine IL-4, IL-8, IL-10, and TNF-α levels correlate with pulmonary function in patients with asthma-chronic obstructive pulmonary disease (COPD) overlap syndrome. Med. Sci. Monit. 22, 2800 (2016).
Article PubMed PubMed Central Google Scholar
-
Chen, J. et al. The elevated CXCL5 levels in circulation are associated with lung function decline in COPD patients and cigarette smoking-induced mouse model of COPD. Ann. Med. 51, 314–329 (2019).
Article PubMed PubMed Central Google Scholar
-
Cota-Gomez, A., Flores, A. C., Ling, X. F., Varella-Garcia, M. & Flores, S. C. HIV-1 Tat increases oxidant burden in the lungs of transgenic mice. Free Radic. Biol. Med. 51, 1697–1707 (2011).
Article PubMed PubMed Central Google Scholar
-
Ensoli, B., Barillari, G., Salahuddin, S. Z., Gallo, R. C. & Wong-Staal, F. Tat protein of HIV-1 stimulates growth of cells derived from Kaposi’s sarcoma lesions of AIDS patients. Nature 345, 84–86 (1990).
Article PubMed Google Scholar
-
Frankel, A. D. & Pabo, C. O. Cellular uptake of the tat protein from human immunodeficiency virus. Cell 55, 1189–1193 (1988).
Article PubMed Google Scholar
-
Ensoli, B. et al. Release, uptake, and effects of extracellular human immunodeficiency virus type 1 Tat protein on cell growth and viral transactivation. J. Virol. 67, 277–287 (1993).
Article PubMed PubMed Central Google Scholar
-
Chang, H. C., Samaniego, F., Nair, B. C., Buonaguro, L. & Ensoli, B. HIV-1 Tat protein exits from cells via a leaderless secretory pathway and binds to extracellular matrix-associated heparan sulfate proteoglycans through its basic region. AIDS 11, 1421–1431 (1997).
Article PubMed Google Scholar
-
Crothers, K. et al. Increased COPD among HIV-positive compared to HIV-negative veterans. Chest 130, 1326–1333 (2006).
Article PubMed Google Scholar
-
Thudium, R. F. et al. Faster lung function decline in people living with HIV despite adequate treatment: a longitudinal matched cohort study. Thorax 78, 535–542 (2023).
Article PubMed Google Scholar
-
Life Span as a Biomarker. The Jackson laboratory https://www.jax.org/research-and-faculty/research-labs/the-harrison-lab/gerontology/life-span-as-a-biomarker.
-
Gibbs, J. et al. An epithelial circadian clock controls pulmonary inflammation and glucocorticoid action. Nat. Med. 20, 919–926 (2014).
Article PubMed PubMed Central Google Scholar
-
Hadden, H., Soldin, S. J. & Massaro, D. Circadian disruption alters mouse lung clock gene expression and lung mechanics. J. Appl. Physiol. 113, 385–392 (2012).
Article PubMed PubMed Central Google Scholar
-
Gan, W. Q., Man, S., Senthilselvan, A. & Sin, D. Association between chronic obstructive pulmonary disease and systemic inflammation: a systematic review and a meta-analysis. Thorax 59, 574–580 (2004).
Article PubMed PubMed Central Google Scholar
-
Taibi, D. M. Sleep disturbances in persons living with HIV. J. Assoc. Nurses AIDS Care 24, S72–S85 (2013).
Article PubMed PubMed Central Google Scholar
-
Asher, G. et al. SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell 134, 317–328 (2008).
Article PubMed Google Scholar
-
Wang, R.-H. et al. Negative reciprocal regulation between Sirt1 and Per2 modulates the circadian clock and aging. Sci. Rep. 6, 28633 (2016).
Article PubMed PubMed Central Google Scholar
-
Thakur, B. K., Chandra, A., Dittrich, T., Welte, K. & Chandra, P. Inhibition of SIRT1 by HIV-1 viral protein Tat results in activation of p53 pathway. Biochem. Biophys. Res. Commun. 424, 245–250 (2012).
Article PubMed Google Scholar
-
Panda, K. et al. HIV-1 tat protein and cigarette smoke mediated ADAM17 upregulation can lead to impaired mucociliary clearance. Cells 13, 2009 (2024).
Article PubMed PubMed Central Google Scholar
-
Spengler, M. L. et al. Core circadian protein CLOCK is a positive regulator of NF-kappaB-mediated transcription. Proc. Natl Acad. Sci. USA 109, E2457–E2465 (2012).
Article PubMed PubMed Central Google Scholar
-
Yang, H. et al. SIRT1 activators suppress inflammatory responses through promotion of p65 deacetylation and inhibition of NF-kappaB activity. PLoS ONE 7, e46364 (2012).
Article PubMed PubMed Central Google Scholar
-
Di Stefano, A. et al. Blood MCP-1 levels are increased in chronic obstructive pulmonary disease patients with prevalent emphysema. Int. J. Chron. Obstruct. Pulmon. Dis. 13, 1691–1700 (2018).
-
Yang, J. et al. Diverse injury pathways induce alveolar epithelial cell CCL2/12, which promotes lung fibrosis. Am. J. Respir. Cell Mol. Biol. 62, 622–632 (2020).
Article PubMed PubMed Central Google Scholar
-
Li, M. et al. Emergence of fibroblasts with a proinflammatory epigenetically altered phenotype in severe hypoxic pulmonary hypertension. J. Immunol. 187, 2711–2722 (2011).
Article PubMed Google Scholar
-
Mulholland, B. S., Forwood, M. R. & Morrison, N. A. Monocyte chemoattractant protein-1 (MCP-1/CCL2) drives activation of bone remodelling and skeletal metastasis. Curr. Osteoporos. Rep. 17, 538–547 (2019).
Article PubMed PubMed Central Google Scholar
-
Xue, W., Ma, J., Li, Y. & Xie, C. Role of CD4+ T and CD8+ T lymphocytes-mediated cellular immunity in pathogenesis of chronic obstructive pulmonary disease. J. Immunol. Res. 2022, 1429213 (2022).
Article PubMed PubMed Central Google Scholar
-
Narendra, D. K. & Hanania, N. A. Targeting IL-5 in COPD. Int. J. Chron. Obstr. Pulm. Dis. 14, 1045–1051 (2019).
Article Google Scholar
-
Zhu, Z. et al. Pulmonary expression of interleukin-13 causes inflammation, mucus hypersecretion, subepithelial fibrosis, physiologic abnormalities, and eotaxin production. J. Clin. Investig. 103, 779–788 (1999).
Article PubMed PubMed Central Google Scholar
-
Aran, D. et al. Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage. Nat. Immunol. 20, 163–172 (2019).
Article PubMed PubMed Central Google Scholar
-
Shafaati, M. et al. The relevance of the circadian timing system role in patients with HIV/AIDS: a quick glance. Int. J. Surg. 109, 2831–2834 (2023).
Article PubMed PubMed Central Google Scholar
-
Maremanda, K. P., Sundar, I. K., Li, D. & Rahman, I. Age-dependent assessment of genes involved in cellular senescence, telomere, and mitochondrial pathways in human lung tissue of smokers, COPD, and IPF: associations with SARS-CoV-2 COVID-19 ACE2-TMPRSS2-Furin-DPP4 axis. Front. Pharmacol. 11, 584637 (2020).
Article PubMed PubMed Central Google Scholar
-
Sundar, I. K. et al. DNA methylation profiling in peripheral lung tissues of smokers and patients with COPD. Clin. Epigenetics 9, 1–18 (2017).
Article Google Scholar
-
Chinnapaiyan, S. et al. TGF-β1 increases viral burden and promotes HIV-1 latency in primary differentiated human bronchial epithelial cells. Sci. Rep. 9, 12552 (2019).
Article PubMed PubMed Central Google Scholar
-
Chinnapaiyan, S. et al. Cigarette smoke promotes HIV infection of primary bronchial epithelium and additively suppresses CFTR function. Sci. Rep. 8, 7984 (2018).
Article PubMed PubMed Central Google Scholar
-
Ramsey, K.M. et al. Circadian measurements of sirtuin biology. Methods Mol Biol. Vol. 1077, 285-302 (Springer, 2013).
-
Faizan, M. I. et al. Genetic diversity leads to differential inflammatory responses to cigarette smoke in mice. Physiol. Rep. 13, e70199 (2025).
Article PubMed PubMed Central Google Scholar
-
Yao, H. et al. SIRT1 protects against emphysema via FOXO3-mediated reduction of premature senescence in mice. J. Clin. Investig. 122, 2032–2045 (2012).
Article PubMed PubMed Central Google Scholar
-
Yao, H. et al. Extracellular superoxide dismutase protects against pulmonary emphysema by attenuating oxidative fragmentation of ECM. Proc. Natl Acad. Sci. USA 107, 15571–15576 (2010).
Article PubMed PubMed Central Google Scholar
-
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 16, 1289–1296 (2019).
Article PubMed PubMed Central Google Scholar
-
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e3529 (2021).
Article PubMed PubMed Central Google Scholar
-
Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888–1902.e1821 (2019).
Article PubMed PubMed Central Google Scholar
-
Sikkema, L. et al. An integrated cell atlas of the lung in health and disease. Nat. Med. 29, 1563–1577 (2023).
Article PubMed PubMed Central Google Scholar
-
Squair, J. W. et al. Confronting false discoveries in single-cell differential expression. Nat. Commun. 12, 5692 (2021).
Article PubMed PubMed Central Google Scholar
Download references
Acknowledgements
The authors would like to acknowledge the funding agencies: NIH grants R01HL147715, R01HL158316 and R01HL167655 awarded to HU and IR. NIH grant R01AI174269 awarded to HU; NIH grant R01HL176254, R03DA057162 and Herbert Wertheim College of Medicine (HWCOM) pilot grant awarded to SC; NIH grants HL60190, HL137282, HL134610 and HL146369 awarded to SMB. We would like to thank Ms Calla Fahey for her technical assistance with IHC staining and Dr. Suvankar Ghorai for his technical assistance with Western Blotting.
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Moses New-Aaron and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Joao Valente. A peer review file is available.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
Reprints and permissions
About this article
Cite this article
Panda, K., Peng, J.Yc., Santiago, M.J. et al. HIV disrupts the lung molecular clock, leading to lung inflammation and features of emphysema. Commun Biol 9, 11 (2026). https://doi.org/10.1038/s42003-025-09284-1
Download citation
This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.
Source: Nature
https://www.nature.com/articles/s42003-025-09284-1
Back to ...
Positively Positive - Living with HIV/AIDS:
HIV/AIDS News
For more HIV and AIDS News visit...
Positively Positive - Living with HIV/AIDS: HIV/AIDS News Archive
|